Schindelhauer and Schwarz (2002) Genome Res. 12: 1815-1826
Supplementary Figure 1
Large version of this figure
Next figures
Back
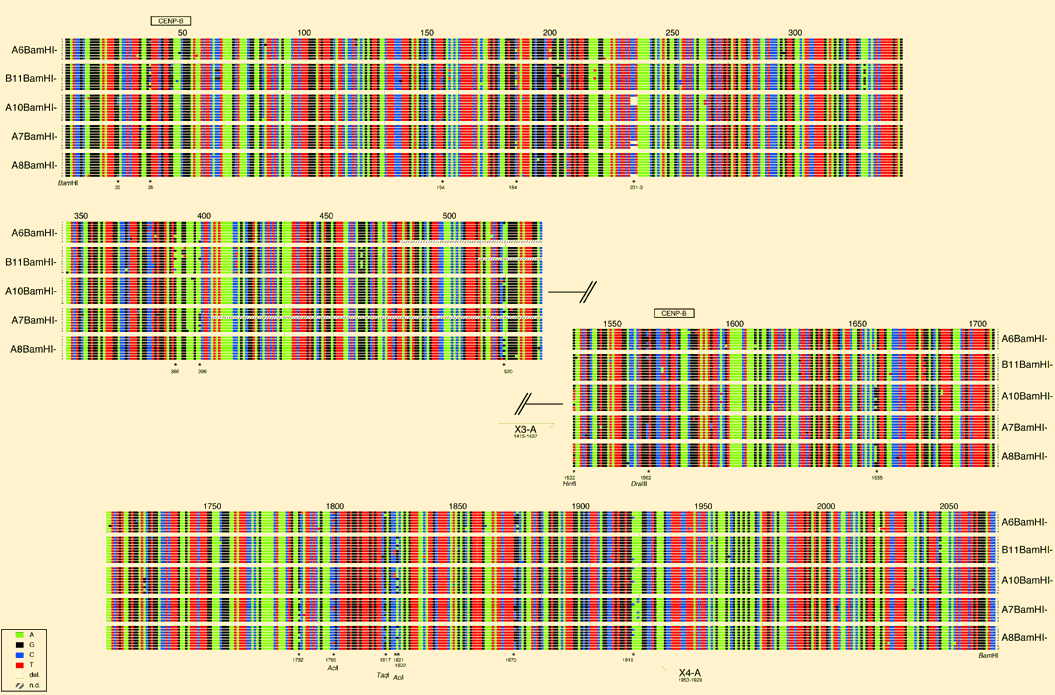
Suppl. figure 1 Four color sequence alignment. Satellite tartan DXZ1 (unordered). Samples of ten 2kb BamHI higher order repeats were subcloned from each of 5 X chromosome alpha satellite PACs and end-sequenced. Subclones from a PAC showing identical end-sequences were excluded because of potential doublet cloning. Identical end sequences of individual subclones A10BamHI-2 and A8BamHI-9 might indicate PAC overlap. In order to compact the sequence information of the 49 kb obtained, little squares were drawn for each nucleotide and painted using the ABI color code (see inset). Sequences are numbered beginning and ending with full BamHI recognition sequences and the gap size between end sequences was guessed. Each row approximates two 0.17 kb monomers. Position of centromere binding protein B-boxes (CENP-B) and primers X3-A and X4-A (Warburton and Willard 1992) used for array specific PCR from total genomic DNA (arrows) are indicated. Positions of a set of "common" variants abundant in more than one PAC sample are marked with asterisks, and restriction sites used to analyse variant fixation are shown. The display illustrates the high overall homogeneity of DXZ1 sequences within and throughout PAC samples, and gives an overview about the distribution of rare and common variants within higher order repeats.
Figure 1 Variant box
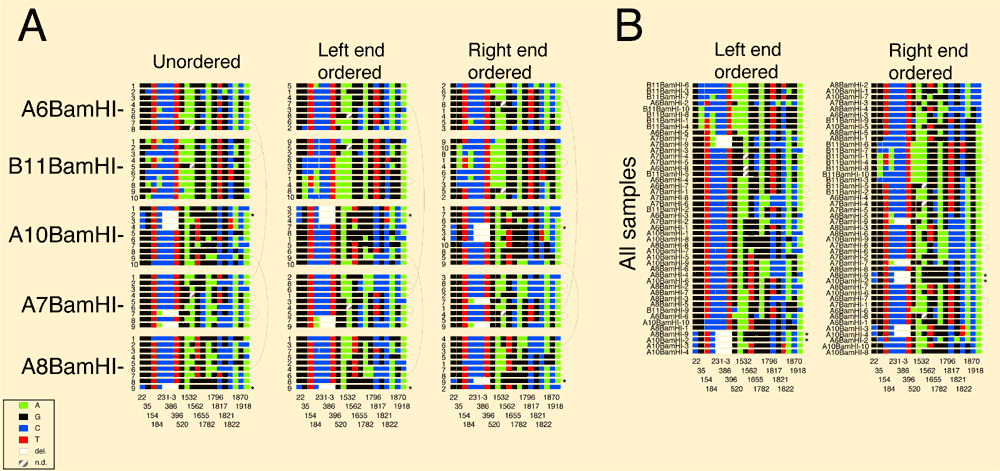
Figure 1 Variant box. For haplotype analysis, the combinations of a set of 18 "common" variants were diagrammed for each of the individual 2kb BamHI subclones from the 5 PAC samples. Little squares were drawn for each nucleotide using the ABI color code (see inset). Variant positions (bottom) belong to the numbering of the satellite sequence in this work (see suppl. figure 1). The 3 bp deletion variant at position 231-233 spans three squares. The variants initially were isolated because they were abundant within a single X chromosome array and typically revealed a fixed transition state within the population. Within the most homogeneous PACs A7, A8, and A10, such common variants account for the majority of the total variation. The slightly lower homogeneity of PACs A6 and B11 is mainly attributable to a higher fraction of regional variants, possibly due to a reduced participation in the overall homogenization process (spatially or temporally). The overall sequence divergence of PACs A6 and B11 could well be considerably higher, because a significant fraction of dispersed BamHI fragments with sizes other than 2kb were not analyzed (Table 2). Of the 3 most homogeneous PACs, only A8 contains such a disruption of the regular BamHI higher order repeat structure (not shown). Ordering the combinations of the 18 common variants derived from PAC samples (A) or from all PACs (B) along their left or right end, neither led to regular patterns reminiscent of a limited number of major haplotypes, nor revealed any obvious relatedness of haplotypes with respect to PAC origin. Of the 46 combinations from individual subclones, 40 are unique and none occurs three times. The six pairs of combinations occuring twice were found within the highly homogeneous PAC A7 (2x), the less homogeneous PAC B11 (containing a duplication of 35-50kb), or between the two homogeneous PACs A10 and A8, or between the homogeneous PAC A7 and diverged PAC A6, as indicated by arches. Asterisks indicate individual clones with identical end-sequences. Several combinations of PAC B11 seem to be more related to each other than to the combinations of other PACs, which to some extent could be explained by a somewhat reduced informativity of the set of 18 variants in this PAC. Interestingly, according to partial restriction mapping, PACs B11, A6 and A7 can not extensively overlap with each other, and all three can not extensively overlap with PACs A8 and A10 (which could overlap almost entirely). Nevertheless, ordering the combinations from all PACs results in many possible relationships within and between the PACs. This analysis suggests a lack of major haplotypes within the array, not supporting homogenization via amplification and replacement. In addition, similar haplotype analyses within narrow regions derived from the 2 or 3 DraIII and non-DraIII clusters of PAC A7 (see suppl. figures 3, 5) suggest that also small regions of amplified haplotypes are very unlikely.
Next figures
Back